Home

01
Join Us, In Saving Lives And Engineering The Future
We are a diverse team of talented people who are dedicated to building technology that radically improves human lives.
February 6, 2018
mvalenzuela
Abstract
Memory failures are frustrating and often the result of ineffective encoding. One approach to improving memory outcomes is through direct modulation of brain activity with electrical stimulation. Previous efforts, however, have reported inconsistent effects when using open-loop stimulation and often target the hippocampus and medial temporal lobes. Here we use a closed-loop system to monitor and decode neural activity from direct brain recordings in humans. We apply targeted stimulation to lateral temporal cortex and report that this stimulation rescues periods of poor memory encoding. This system also improves later recall, revealing that the lateral temporal cortex is a reliable target for memory enhancement. Taken together, our results suggest that such systems may provide a therapeutic approach for treating memory dysfunction.
Introduction
Research on human episodic memory has shown that whether information is remembered or forgotten depends on neural events that transpire during encoding. Spectral power recorded using intracranial electrophysiology1 and blood-oxygen-level-dependent functional magnetic resonance imaging (fMRI) signal2 show that activity in many cortical and subcortical regions differentiates learned information that is likely to be remembered from information that is likely to be forgotten. Differences in neural activity during encoding therefore predict intra-individual variability in later memory performance, suggesting that modulating neural activity when the brain is unlikely to encode successfully could improve overall performance by rescuing network activity.
A promising tool for modulation of neural activity is direct brain stimulation, in which electrical current is applied via electrodes implanted on or directly in the brain parenchyma. Direct brain stimulation is a standard tool in the treatment of motor dysfunction in Parkinson’s disease and seizure onset in epilepsy3,4,5,6, and has recently been explored as a therapy for psychiatric conditions7,8. Direct brain stimulation treatment commonly involves continuous (i.e., open-loop) high-frequency stimulation, although recent work has suggested improved effectiveness when applying stimulation in response to specific brain states (i.e., closed-loop9,10).
Several studies have used direct brain stimulation of the human hippocampus and medial temporal lobes (MTLs), core regions of the brain’s memory network, to modulate neural activity and performance during memory tasks. These studies used open-loop designs, meaning that stimulation was not delivered in response to ongoing neural activity. Although some studies using this approach have suggested that such stimulation improves memory11,12,13,14, others have failed to show improvements or have shown disruption15,16,17,18,19,20. This literature shows that direct open-loop stimulation of the hippocampus and MTL is unlikely to reliably improve memory, and suggests that stimulating other regions in the memory network may be more effective for memory enhancement.
Encoding activity in the left lateral temporal cortex (including the middle portions of the inferior, middle, and superior temporal gyri) measured with fMRI2 and intracranial electroencephalography (iEEG)1,21,22 predicts memory performance, and stimulation mapping of this area has been shown to evoke memory-like recollective phenomena23,24. Prior invasive25,26,27 and non-invasive28,29,30stimulation studies also suggest this region to be a prime target for memory modulation. However, these studies did not characterize the effect of lateral temporal cortex stimulation in comparison with other targets. The previous work also did not use stimulation to rescue intervals of poor memory encoding by timing stimulation based on recordings of ongoing neural activity.
Here we evaluate the use of lateral temporal cortex as a stimulation target by deploying a closed-loop architecture for sensing and stimulating the brain during a memory task. We compare the effect of lateral temporal cortex stimulation to both a within-subject non-stimulated condition and an independent non-lateral temporal stimulation group. We use multivariate classifiers that are individualized to each subject to decode neural activity during memory encoding and trigger stimulation online in response to patterns of neural activity that are associated with later forgetting. By using classification within a closed-loop system, we maximize sensitivity to detect poor encoding states and account for the fact that direct brain stimulation has distributed effects on physiology31,32 that depend on the state of the brain at the time of stimulation delivery33,34,35,36,37.
Results
Classifying recall probability from neural activity
We recruited 25 neurosurgical patients undergoing clinical monitoring for epilepsy to participate in sessions of a delayed free recall memory task (Fig. 1a). Subjects performed at least three record-only sessions of free recall from which we trained a multivariate classifier to discriminate patterns of neural activity during encoding that predicted memory. We fit penalized logistic regression classifiers to record-only data from each subject, producing a set of model weights that map features of iEEG activity to an output probability of later word recall (Fig. 1b).
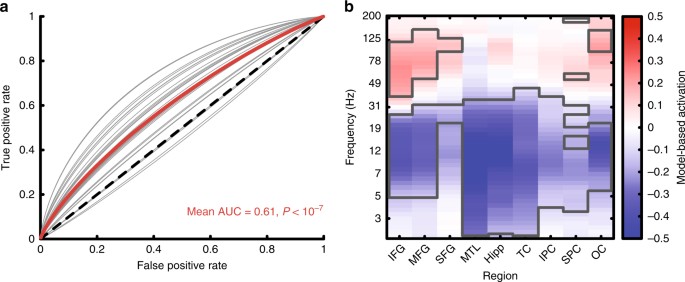
We then used this model in subsequent sessions to decode the probability of recall from neural activity online during the encoding phase of the task (Fig. 1c). On Stim lists, if the predicted probability of recall was below 0.5, the system triggered 500 ms of bipolar stimulation across an adjacent pair of channels. We predicted that this would disrupt poor encoding states and improve memory performance. On NoStim lists, we used the classifier to estimate recall probabilities but did not trigger stimulation. This allowed us to control for the memory state when assessing the effect of stimulation on recall probability of stimulated items. It also allowed us to determine whether the record-only classifier generalized to the new closed-loop session. We found that these classifiers reliably discriminated recalled from not recalled words in the new session (NoStim list area under the curve (AUC) = 0.61, t(28) = 7.5, P < 10−7 by a one sample t-test, Fig. 2a). To determine what information the model used to classify encoding trials we computed a Forward Model for each patient using the classifier weights and neural encoding activity34,38. Across subjects, this showed that the classifiers relied on increased high-frequency activity (HFA) across the brain (frontal, temporal, parietal, and occipital cortex), along with widespread decreases in low-frequency activity to predict words likely to be recalled (Fig. 2b; clusters outlined in gray were significant at the P < 0.05 level by a one-sample t-test).
Stimulation increased the probability of word recall
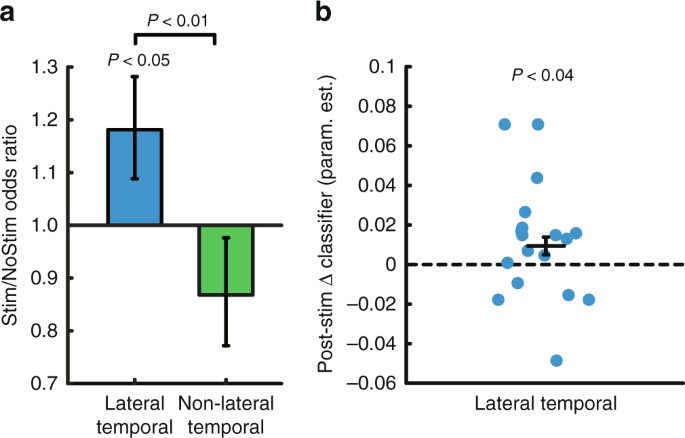
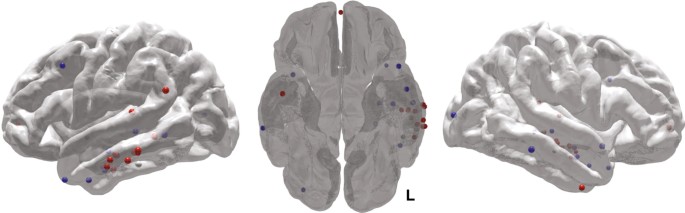
We used a generalized linear mixed effects (GLME) model (logistic regression with a binomial distribution) to model the individual trial-level recalled/not recalled data across subjects, to determine how stimulation affected the probability of word recall. The linear mixed effects approach allows us to estimate the effect of stimulation on probability of recall while accounting for heterogeneity in the amount of data collected from each subject. The resulting parameter estimates reflect the change in odds associated with each condition (Stim/NoStim and Lateral temporal cortex/Non-lateral temporal). Stimulation of lateral temporal cortex increased the odds of recalling stimulated words compared with matched non-stimulated words (odds ratio = 1.18, t(3,828) = 2.04, P = 0.04, Fig. 3a) and compared with Non-lateral temporal stimulation (t(5,864) = 2.60, P = 0.009). Stimulation led to a nonsignificant decrease in recall odds in the Non-lateral temporal group (odds ratio = 0.87, t(2,036) = −1.20, P = 0.23). The stimulation targets that led to increased memory performance in the lateral temporal cortex group clustered in the middle portion of the left middle temporal gyrus (Fig. 4). Using a log-binomial model39 to estimate the relative change in recall probability, we found that lateral temporal cortex stimulation increased the relative probability of item recall by 15% (t(3828) = 2.31, P = 0.02, Supplementary Table 1). Subjects stimulated in lateral temporal cortex were also more likely to remember unstimulated items that flanked the stimulation (t(960) = 2.46, P = 0.01; interaction with Non-lateral temporal stimulation t(1,406) = 2.48, P = 0.01).
We observed these different stimulation-related memory outcomes in spite of the fact that the groups were matched in several ways. The proportion of words on which the closed-loop classifier triggered stimulation was roughly 0.50 (as intended) and did not differ between the lateral temporal cortex and Non-lateral temporal groups (lateral temporal cortex = 0.52, Non-lateral temporal = 0.50, t(5,698) = 0.30, P = 0.71 by linear mixed effects model). Stimulation also did not differ between the groups as a function of word position within the list (position × group interaction t(5,696) = − 0.84, P = 0.40). The groups showed similar memory performance during the record-only sessions (lateral temporal cortex = 26.1%, Non-lateral temporal = 29.8%, t(24) = −0.87, P = 0.39 by two-sample t-test). We also analyzed subsequent memory effects (SMEs) at the stimulated electrodes in the record-only data, to determine whether the stimulated electrodes showed different contributions to memory performance across the groups. We found that power across the frequency spectrum (2–200 Hz) was similarly predictive of memory for the lateral temporal cortex and Non-lateral temporal groups (Supplementary Figure 1a, b), indicating no difference between groups in the stimulated electrode’s involvement in encoding processes. We also did not find any relation between the size of the SME at the stimulated electrode and stimulation’s effect on behavior (Supplementary Figure 1c).
Neural evidence for improved encoding following stimulation
We next asked how lateral temporal cortex stimulation influenced neural activity to support memory encoding by comparing HFA (70–200 Hz) between Stim and NoStim conditions. High-frequency power predicts memory success1,37,40,41 and is thought to reflect excitation of neural populations engaged during cognition42,43. When comparing evoked HFA following stimulation to matched unstimulated periods in electrodes placed in the same lobe as the stimulation target, we did not observe significant increases in HFA power following lateral temporal cortex stimulation (Supplementary Fig. 2). This suggests that lateral temporal cortex stimulation did not reliably modulate activity in these target regions when aggregating data across subjects, which was not surprising given that stimulation has heterogeneous effects on downstream targets31,32, and that our subjects each possessed a unique recording montage. We reasoned that a more sensitive approach would be to assess the effect of stimulation on neural activity using a model that accounts for this between-subject variability.
The classifier that we trained on each subject’s record-only data is an individualized model relating neural activity in that person to a probability of recall, so we next asked whether these classifiers could be used to decode the effects of stimulation on neural activity. We assessed this by computing the change in whole-brain classifier output for consecutive words (wi + 1 and w i , for all stimulated words i). We did the same for matched words on NoStim lists and compared these ∆ classifier values between the Stim and NoStim conditions for the lateral temporal cortex group using a mixed model. This analysis showed increases in classifier output following stimulation compared to the control non-stimulated condition (estimate = 0.01, t(1,404) = 2.06, P = 0.04 by linear mixed effects model, Fig. 3b), suggesting that stimulation’s influence on neural activity was evident when using the whole-brain classifier to decode the change in neural activity post- stimulation.
The original publication can be found here.